Shaozheng Song1,2,
Zhengyi He1,
Junyan Mei1,
Zhengqiang Qi1,
Si Chen1,
Sheng Xu1,
Yugu Yuan1,2,
Yong Cheng1,2 ![]() ,
,
For correspondence:- Yong Cheng Email: ssz0610@126.com Tel:+8615906181002
Received: 5 January 2015 Accepted: 6 April 2016 Published: 27 May 2016
Citation: Song S, He Z, Mei J, Qi Z, Chen S, Xu S, et al. Affinity purification of recombinant human plasminogen activator from transgenic rabbit milk using a novel polyol-responsive monoclonal antibody. Trop J Pharm Res 2016; 15(5):905-911 doi: 10.4314/tjpr.v15i5.2
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To develop processes for effective isolation and purification of recombinant human plasminogen activator (rhPA) from transgenic rabbit milk.
Methods: Immunoaffinity chromatography was selected and improved by a special polyol-responsive monoclonal antibody (PR-mAb). Alteplase was used as immunogen because of its similarity to rhPA in terms of structure. The PR-mAb was prepared by hybridoma technology and screened by ELISA-elution assay. Screening antibody was performed using rhPA milk in an ELISA-elution assay. The antibody clone C4-PR-mAb was selected for immunoaffinity chromatography. The rhPA was effectively bound to immobilized C4-PR-mAb on the column and was eluted with Tris buffer comprising 0.75 mol/L ammonium sulfate and 40n% propanediol (pH7.9). The rhPA was further purified by passing through Chromdex75 gel filtration column.
Results: There were 12 hybridoma strains selected into the polyol-responsive mAbs screen step and three hybridoma strains were superior for producing PR-mAbs (C1, C4, C8). The rhPA can be purified from transgenic rabbit milk and maintained a higher thrombolytic activity in vitro by FAPA.
Conclusion: The results demonstrate the suitability of the alternative approach used in this study. Using immunoaffinity chromatography and gel filtration column is feasible and convenient for extracting rhPA from milk, and should be useful for purifying other tPA mutants or other novel recombinant milk-derived proteins.
Introduction
Natural human tissue plasminogen activator (tPA) is secreted by vascular endothelial cells displaying thrombolytic activity. Currently on the market, clinical tPA is used for treating acute myocardial infarction, brain death, pulmonary embolism, deep venous thrombosis, and other thrombotic diseases and is produced by prokaryotic or eukaryotic cell expression system (e.g., CHO) [1-3]. Some modifications on molecular structure have resulted in recombinant tPA (rtPA or rhPA) with better bioactive characters, such as higher specificity to thrombus, longer half-life in blood, and less amount of bleeding [1,2]. However, commonly used processes for rhPA are based on prokaryotic or eukaryotic vector which impose many limitations on rhPA development, i.e., its technology and cost [1,4].
To increase the half-life and thrombolytic activity of tPA-type drugs, a novel tPA mutant (rhPA) was designed by our research team. With only K2 and P structure domain, it was different from Alteplase, which contains F, E, K1, K2, and P domains. It was produced by using a transgenic rabbit mammary gland bioreactor, which may be a popular approach for expressing recombinant foreign protein more safely. This bioreactor also produces higher output than prokaryotic or eukaryotic cell expression system. If its purity is further improved, and its activity is preserved in the manufacturing process, rhPA has great potential as a new application in thrombolytic medicine. However, effective purification for this novel recombinant protein is needed.
Many methods of protein extraction from milk have been studied [5,6], among which immuno affinity chromatography(IAC) is a convenient and efficient approach because of its simple one-step operation and high specificity to target protein [7,8]. However, a strongly acidic buffer (pH is less than or equal to 3.0) is often used to elute and dissociate the target protein, which was not beneficial in protecting the molecular conformation and bioactivity of rhPA.
In order to preserve the functional domains, it would be best to assemble an affinity column that allows protein dissociation from immobilized ligand. Polyol-responsive monoclonal antibody (PR-mAb) is a kind of special ligand that can release the target protein from the IAC column gently and easily [8]. The application of PR-mAbs to the affinity column would improve the defects because of its slightly alkaline elution buffer instead of strong acidic condition, consisting of Tris buffer with ammonium sulfate and propylene glycol (pH7.9) which have little damage to protein stability [9].
Preparation of specific antibodies is very important in affinity technology; finding a suitable immunogen is a key link. Alteplase, a commercial drug, can be used as immunogen for anti-rhPA antibodies, which has a structure similar to that of rhPA and can be obtained with high purity (> 99 %).However, to prepare specific antibody for rhPA, Alteplase was used as the immunogen. After conducting the Elisa-elution screening method, three strains of PR-mAbs were successfully obtained.
Methods
This study was approved by the Institutional Animal Care and Use Committee of Yangzhou University (China) and performed according to the Guide for the care and use of laboratory animals [10]. Chemicals were purchased from Sigma Chemical Co. and China unless otherwise specified.
Immunization and hybridoma cell culture
Six-week old BALB/c mice were immunized by Alteplase (purchased from Subei People's Hospital of Jiangsu province, China). QuickAntibodyTM immunologic adjuvant was used to enhance the immune response of Alteplase, mixed with Alteplase of 1:1 volume. Each mouse was injected intramuscularly with 5 μg mixture of Alteplase-QuickAntibody twice, at an interval of 21 days. At 14 days after the secondary immunization, booster injection was given intramuscularly with Alteplase-PBS solution at 10 μg per mouse. Three days after booster injection, the mouse that presented higher serum level of antibodies was selected for cell fusion.
Electro fusion technique was used in cell fusion. However, spleen cells and myeloma cells were mixed at a ratio of 1:1 and fused in electro fusion solution (0.3 M mannitol, 0.1 mM CaCl2, and 0.1 mM MgCl2) using cell fusion generator. The distance between the two electrodes was 2 mm. Membrane fusion was performed by three electrical pulses; first with alternating current at 40 V for 30 s and then with two times of direct current at 325 V (162.5 V/mm) for 30 μsat an interval time of 0.5 s. The fused cells were centrifuged (800 g, 5min) and the cell pellets were suspended with HAT/DMEM medium and planted into 96-well platespre-coated with feeder cells.
Screening for anti-rhPA mAbs
Indirect ELISA was used here. The rhPA milk whey of transgenic rabbit was immobilized to ELISA plate as the detecting antigen. In another ELISA plate, the normal rabbit milk whey was immobilized as control antigen to exclude the false positive results that would be caused by the nonspecific binding of antibodies to other components from rabbit whey. The supernatant of each cell culture was the primary antibody, SP20 cell culture acted as negative control, and PBS acted as blank control. After three times subcloning by limited dilution assay, positive hybridoma could be determined.
Screening for PR-mAbs
ELISA-elution assay was performed as described previously [12]. Here, the rhPA milk whey was immobilized, cross-reacting with the primary antibody (supernatant of positive hybridoma cells) for 2 h at 37 °C. SP20 cell culture acted as negative control, and PBS acted as blank control. After washing, Tris buffer containing 0.75 M ammonium sulfate and 40 % (v/v) propylene glycol (pH 7.9) was added to each sampling hole and kept at room temperature (24 °C) for 20 min to dissociate the binding of antibody to antigen. The secondary antibody (HRP conjugated goat anti-mouse IgG antibody) was diluted with dilution buffer (PBS with 1 % FBS) at a volume ratio of 1:2000. After color development, each well was read by microplate reader at 450 nm.
Large scale preparation and purification for PR-mAb
Eight-week old multiparous BALB/c mice were intraperitoneally injected with 0.5 mL liquid paraffin,followed by the PR-mAb hybridoma cells of 5 × 106 cells per mouse 1 week later. After 5 days, peritoneal fluid was collected and purified by ProteinA affinity column.
Preparation for immunoaffinity column
The purified antibody was dialyzed with 3.5 KDa molecular weight cutoff dialytic-bags against coupling buffer (0.2 M NaHCO3, 0.5 M NaCl) overnight at 4 °C and concentrated via PEG20000. This antibody solution was transferred to the CNBr activated Sepharose for conjugation. The process was performed at room temperature (24 °C) overnight, whereas the uncoupled CNBr groups were blocked in the blocking buffer (20 mM Tris-HCl, pH 8.3) for 4 h at the same temperature. Before the resin could be used, 0.05 M Tris-HCl (pH 8.5) and 0.2 M acetate (pH 4.6) were used to wash the mAb-Sepharosesix times alternately. Finally, precooled PBS was transferred to suspend the resin.
Purification of rhPA using the immunoaffinity column
Before application to the IAC, milk sample was processed as follows rabbit milk was centrifuged (8,000 rpm) at 4 °C for 30 min and the supernatant of whey was separated from transgenic rabbit milk; 35 % saturated ammonium sulfate was added to the supernatant for precipitating the rhPA component and then the pellet was dialyzed with 3.5 KDa molecular weight cutoff dialytic-bags against PBS overnight at 4 °C.
After pretreatment on milk, the crude sample was run on the C4-PR-mAb coupled column. AKTA explorer system with UV detector (280 nm), sample loops, fraction collector and analysis software was used in the IAC. The packed column was washed and equilibrated with PBS at 1 mL/min. The crude rhPA solution was loaded into the column at a flow speed of 1 mL/min and, PBS used to elute the unbinding proteins out of the affinity column. When the flow through the baseline was stable, elution buffer (TE buffer with 0.75 mol/L ammonium sulfate and 40 % propanediol, pH 7.9) was run on the IAC at 1 mL/min to dissociate the rhPA from the mAb-sepharose. The peak fractions were collected and analyzed by SDS-PAGE.
Further purification of rhPA using gel filtration chromatography
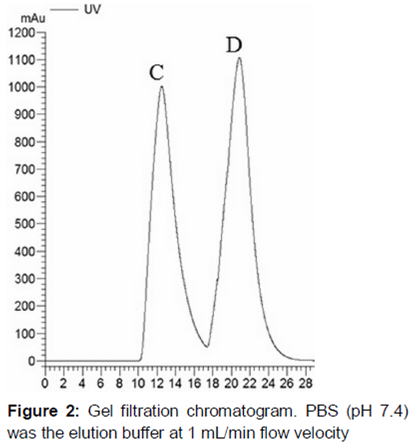
Chromdex 75 prep grade gel filtration was used to remove the other large proteins. IAC processed sample was run on the gel filtration column and eluted with PBS (pH 7.4) at 1 mL/min. The other proteins with larger molecular weight passed through the column first, and the rhPA fraction followed.
Purity, antibody cross-reactivity and thrombolytic activity of the purified rhPA
SDS-PAGE was used to analyze the purity of rhPA product from immunoaffinity step and gel filtration step. The unstained gel was applied in Western blot analysis to identify the rhPA. However, the gel was transferred to the 0.45 micrometers PVDF membrane in transfer buffer at 1 mA/cm2 for 3.5 h, Furthermore, the membrane was placed in blocking buffer at 4 °C overnight. The primary antibody was the purified PR-mAb, which was diluted at 1:5000 in PBS and incubated with membrane at 37 °C for 1.5 h. The secondary antibody conjugated with HRP was added and held at 37 °C for 1.5 h. After washing thrice with washing buffer, DAB substrate solution was used for blot development, and incubation was done for 20 min at room temperature (24 °C).
ELISA was used to determine the antibody cross-reactivity of each fraction collected from the chromatography process. Procedure for this test was similar to the antibody screening section. However, the coating antigen was replaced with each fraction of chromatography, and the primary antibody was replaced with the purified PR-mAb. The other operations were the same as the part of antibody screen.
Fibrin agarose plate assay (FAPA) was carried out to evaluate the thrombolytic activity of the purified rhPA. Fibrin-thrombin-agarose gel (100 mL) was composed of 1 g agarose, 1 g fibrinogen, and 100 U thrombin dissolved in PBS. Agarose was brought to a boil, and after cooling to 45 °C, boiled agarose was immediately mixed with fibrinogen and thrombin (37 °C). The warm mixture was then dispensed into Plexiglass dishes. When the solution cooled to room temperature, fibrin-thrombin-agarose solidified to a gel state. Sample wells were drilled in each gel and filled with 20 μL sample solution. In thrombolytic step, each dish was incubated at 37 °C overnight. The activity was determined by the diameter size of the thrombin-dissolving ring.
Statistical analysis
Data analysis was performed by descriptive analysis and standard curve using SPSS 21 (SPSS Institute, Cary, NC, USA).
Results
Preparation of anti-rhPA monoclonal antibody
QuickAntibodyTM adjuvant was used in mouse immunization. After boost immunization, the serum titer reached 1:1010. In this study, the dose of immunogen was reduced to 20 μg per mouse, and the experimental days were shortened to 35 days. Improvements in immune dose and period were observed with the above mentioned procedure compared with the common Freund's adjuvant procedure.
A total of 814 hybridoma clones appeared in a96-well plate. Standard ELISA was used to screen positive cell strains producing antibodies that could react with the coating antigen, rabbit milk, which contained rhPA. After three times of subcloning, 12 hybridoma strains were selected for the polyol-responsive mAbs screen step.
Screening of polyol-responsive monoclonal antibody
The 12 positive hybridomas detected in standard ELISA were rescreened here. Comparing the OD value of the two kinds of ELISA, the wells with a significantly decrease in OD were considered to contain polyol responsive mAbs. According to Table1, three hybridoma strains were superior in terms of producing PR-mAbs, as follows: C1, C4, and C8. The values of D-OD reached 5 0 % [10,12].
C4 hybridoma strain was selected to prepare the PR-mAb for immunoaffinity ligand. ProteinA column was used to purify the C4 PR-mAb from mouse ascites.
Immunoaffinity purity of rhPA in rabbit milk
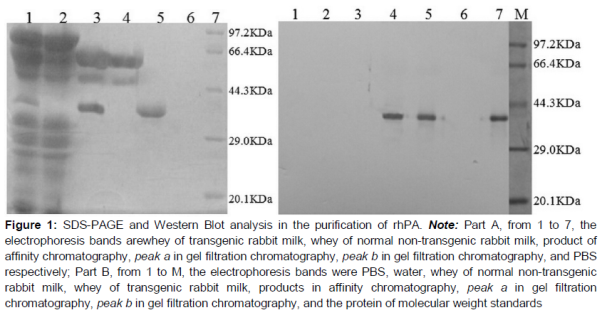
SDS-PAGE gel showed the sample with other two bands at 55–70KDa, which were not the molecular weight of rhPA (). This result suggested that the fraction contained proteins, but not rhPA.
Therefore, gel filtration chromatography was applied to separate the other unrelated proteins. According to the principle of molecular sieve, the large protein was first passed through the column. rhPA, which had smaller molecular weight was considered in the second peak b (). SDS-PAGE showed the peak b fraction only with a single band, and the band was allocated at 39 KDa, which was the molecular weight of rhPA (). Further determination of crosslink activity for the fraction of peak b was performed by Western Blot (). The results suggested that the target protein rhPA was contained in the second peak b.
Activity of purified rhPA
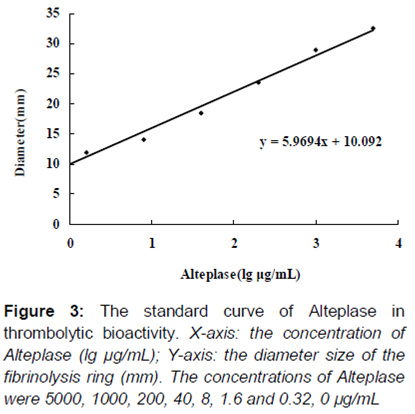
FAPA was applied to evaluate the thrombolytic bioactivity of purified rhPA in vitro, and the index was the diameter size of the fibrinolysis ring. As shown in , well 10 contained the purified rhPA at a concentration of 1 μg/mL, and the thrombolytic activity was presented with 24.0 mm diameter of the fibrinolysis ring. The purified rhPA (1 μg/mL concentration) was presented with 24.0 mm diameter of the fibrinolysis ring of thrombolytic activity. A standard curve () was drawn using the concentrations of Alteplase, as follows: 5000, 1000, 200, 40, 8, 1.6, 0.32 and 0 µg/mL. To reach the same thrombolytic effect of rhPA, the concentration of Alteplase was supposed to be 214 µg/mL, which was much higher than the dose of purified rhPA. Thus, this purified rhPA displayed a 214-fold increase in activity to dissolve thrombus, compared with Alteplase by FAPA.
Discussion
rhPA is our originally designed mutant of tPA. This recombinant protein was produced by a bioreactor, rabbit mammary gland, which has advantages in natural molecular structure, protein amount, and safety. However, purification of the recombinant protein from milk is not an easy process [13]. Immunoaffinity chromate-graphy is widely adopted, but the commonly used elution buffers often contain an acid agent or a chaotropic reagent, which would reduce the activity of target protein [14-16]. As for rhPA, its bioactivity relies on its protein structure. Therefore, a gentle method for purifying rhPA is necessary. In this study, immunoaffinity chromatography coupled with polyol responsive mAb was used. PR-mAbs as immunoaffinity ligands have been reported, and these presented good performance on RNA polymerase and RNA polymerase II [9-11,15]. However, these have not been used in the extraction for rabbit milk protein. This paper described the preparation for PR-mAbs and the application of the C4-PR-mAb in immunoaffinity chromatography for rhPA purification.
Three steps were required in this purification research. The first step involved extracting crude sample from rabbit milk, using salting out, ultrafiltration, and dialysis. The second step included immunoaffinity chromatography with C4-PR-mAb; rhPA and rhPA purity were not high. The other two bands appeared obviously (A). Thus, we initially suspected that C4-PR-mAb crosslinked with other proteins and not only rhPA. However, the normal rabbit milk could not cross-react with the serum antibody of the immunized group, and this was also impossible for C4-PR-mAb (25 and 50 KDa). Therefore, if our first hypothesis was true, it would explain the following: except for rhPA, another novel protein was expressed by the transgenic rabbit. However, this explanation did not have a reliable basis. A was worth mentioning again. The band at 64 KDa may have resulted from the combination of rhPA and the light chain of the antibody (39 KDa + 25 KDa = 64 KDa). The band at 50 KDa may be the heavy chain of antibody (50 KDa). This is not a coincidence because antibody ligand falling off column in affinity chromatography often occurred. Thus, in B, the two fractions with negative reactivity for C4-PR-mAb may be related to the rhPA epitopes covered by the C4-PR-mAb ligand, which was detached from the column. rhPA with a single molecular was our objective. Thus, through Chromdex 75 prep grad gel filtration chromatography, these two bands were successfully removed, and a pure rhPA was ultimately obtained.
Conclusion
C4-PR-mAb is the key factor in this immunoaffinity purification for rhPA. rhPA is a novel recombinant mutant protein without F, E, and K1 structure domain, but the structure is similar to that of Alteplase. Based on this point, Alteplase is creatively selected as immunogen. The rhPA can be purified from transgenic rabbit milk through the above-described method, and it displayed 214-fold higher activity in dissolving thrombus than Alteplase. This process for antibody and purification development is our original design and helps us to determine rhPA PR-mAbs. Meanwhile, we successfully purified rhPA from milk. This method would be useful for other studies on similar novel recombinant proteins.
Declarations
Acknowledgement
References
Archives


News Updates